Уровни регуляции вегетативных функций. Гипоталамус как высший подкорковый центр регуляции вегетативных функций. Подкорковые функции Понятие о нервном центре. Основные свойства нервных центров
Функции промежуточного мозга
Благодаря расположению в промежуточном мозге многих вегетативных ядер, эндокринных желез (гипофиз, эпифиз), зрительного бугра, он:
1) является высшим подкорковым центром вегетативной нервной системы, обеспечивающим выполнение вегетативных функций, связанных с гомеостазом и обменными процессами (белковым, жировым, углеводным, водно-солевым), терморегуляцией;
2) обеспечивает интеграцию всех видов чувствительности организма, заключающуюся в сопоставлении информации, поступающей по различным каналам связей, и оценке её биологической ценности;
3) обусловливает эмоциональное поведение, связанное с мимикой, жестами, изменениями в функции внутренних органов;
4) выполняет гуморальную регуляцию посредством гормонов, выделяемых гипофизом и эпифизом.
Анатомия
Промежуточный мозг сзади граничит со средним мозгом, а спереди - с конечным мозгом. Граница с ним проходит с до реальной поверхности по мозговым полоскам зрительных бугров, с вентральной - впереди зрительного перекреста. В промежуточном мозге различают следующие отделы: тапамическую область, гипоталамус и III желудочек.
К таламической области относятся таламус, метаталамус и эпиталамус.
Таламус (задний таламус, зрительный бугор), t halamus d o rsalis, имеет овоидную форму. Передний отдел заострён и называется передним бугорком, tuberculum anterius thalami, задний утолщён и называется подушкой, pulvinar. Медиальные поверхности зрительных бугров образуют боковые стенки III желудочка, а верхние поверхности участвуют в образовании дна центральной части боковых желудочков. Медиальные поверхности соединяются непостоянным межталамическим сращением, adhesio interthalamica, и отделяются от верхних поверхностей мозговой полоской таламуса, stria medullaris thalamica. Боковые отделы таламуса прилежат к хвостатому ядру конечного мозга, отделяясь от него пограничной бороздой, sulcus terminalis.
Таламус состоит из серого вещества, представленного более чем 40 ядрами. Наиболее крупными и функционально более важными из них являются передние, медиальные и латеральные. Таламус является подкорковым центром всех видов чувствительности, кроме обонятельной, вкусовой и слуховой. В его ядрах происходит переключение на последний, как правило, на 3-й нейрон всех афферентных путей.
Под таламусом находится субталамическая область, regio subthalamica, которая вентрально переходит в покрышку ножек мозга. К этой области относится небольшой участок мозгового вещества, в который из среднего мозга продолжаются и в котором заканчиваются красное ядро и чёрное вещество. Сбоку от чёрного вещества располагается субталамическое ядро (люисово тело), nucleus subthalamicus.
Таламус имеет множественные связи с корой больших полушарий, со стриопаллидарной системой, средним мозгом, мозжечком, подталамической областью.

Метаталамус (заталамическая, забугорная область), meta th alam u s, представлен парными медиальными и латеральными коленчатыми телами. Латеральное коленчатое тело, corpus geniculatum laterale, находится сбоку от подушки зрительного бугра. В него вступают волокна зрительного тракта. Посредством ручек верхних холмиков латеральные коленчатые тела соединяются с верхними холмиками.
Под подушкой несколько внутри и кзади от латеральных коленчатых тел располагаются медиальные коленчатые тела, corpus geniculatum mediale, которые посредством ручек соединяются с нижними холмиками. В клетках этих коленчатых тел заканчиваются волокна латеральной слуховой петли. Как уже отмечалось, латеральные коленчатые тела и верхние холмики среднего мозга являются подкорковыми центрами зрения, а медиальные коленчатые тела и нижние холмики - подкорковыми центрами слуха.
Эпиталамус (надталамическан, надбугорная область), e pith alamus, состоит из 5 небольших образований. Самым крупным из них является шишковидное тело (шишковидная железа, эпифиз мозга), corpus pineale (glandula pinealis, epiphysis cerebri), весом 0,2 г. Название дано за сходство по форме с шишкой сосны (pinus, сосна). Оно располагается в бороздке между верхними холмиками среднего мозга. Посредством поводков, habenulae, эпиталамус соединяется со зрительными буграми. В этих местах имеются расширения - это треугольник поводка, trigonum habenulae. Части поводков, входящие в эпиталамус, образуют спайку поводков, comissura habenularum. Снизу от эпиталамуса имеются поперечно расположенные волокна эпиталамическая спайка, comissura epithalamica. Между этой спайкой и спайкой поводков внутрь эпиталамуса вдаётся шишковидное углубление, recessus pinealis.
Гипоталамус (подталамическая, подбугорная область), hypothalamus , включает нижние отделы промежуточного мозга; зрительный перекрест, зрительные тракты, серый бугор, воронку, гипофиз и сосцевидные тела.
Зрительный перекрест, chiasma opticum, образован медиальными волокнами nn. optici, которые переходят на противоположную сторону и вступают в состав зрительных трактов, tractus opticus. Тракты располагаются медиальнее и кзади от переднего продырявленного вещества, огибают ножку мозга с латеральной стороны и двумя корешками вступают в подкорковые центры зрения: латеральный корешок вступает в латеральное коленчатое тело, а медиальный в верхний холмик крыши среднего мозга.
Серый бугор, tuber cinereum, находится сзади зрительного перекреста. Нижняя часть бугра имеет вид воронки, infundibulum, на которой подвешен гипофиз. В сером бугре локализуются вегетативные ядра.
Гипофиз, hypophysis, располагается в sella turcica тела клиновидной кости, имеет бобовидную форму и вес 0,5 г. Гипофиз, как и эпифиз, относится к железам внутренней секреции.
Сосцевидные тела, corpora mammillaria, сосредоточены между серым бугром и задним продырявленным веществом. Они белого цвета, имеют сферическую форму, их диаметр - около 0,5 см. Внутри сосцевидных тел находятся подкорковые ядра (центры) анализатора обоняния.
В гипоталамусе имеется более 30 ядер, которые разделены по локализации на три области - переднюю, промежуточную и заднюю. Нейроны многих ядер вырабатывают нейросекрет, который по отросткам нейронов транспортируется в гипофиз. Данные ядра называются нейросекреторными. Наиболее крупными и нейросекреторными ядрами в передней области гипоталамуса являются супраоптическое (надзрительное) и паравентрикулярное; в промежуточной зоне-нижне- и верхнемедиальные, дорсальное, серобугорное ядро и ядро воронки; в задней области - медиальное и латеральное ядра сосцевидного тела и заднее гипотапамическое ядро. Все упомянутые ядра относятся к высшим вегетативным ребрам и имеют широкие нервные и гуморальные связи с гипофизом, что дало основание объединить их в гипоталамо-гипофизарную систему.
III желудочек, ventriculus tertius, занимает центральное положение в промежуточном мозге, имеет вид щели, ограничивается шестью стенками, которые формируются определёнными образованиями.
Стенки желудочка выстланы эпендимой. Сосудистая основа верхней стенки является производной сосудистой оболочки головного мозга и состоит из двух листков (дупликатуры) этой оболочки. Между листками сосудистой основы располагаются две внутренние мозговые вены, vv. cerebri internae, которые сливаются в большую мозговую вену (вену Галена), v. cerebri magna, обеспечивающую отток крови в прямой синус. Ворсинки нижнего листка сосудистой основы образуют сосудистое сплетение, plexus choroideus. Это сплетение из полости желудочка покрыто эпителиальной пластинкой lamina epitelialis, производной эпендимы.
Полость III желудочка сообщается с IV желудочком посредством водопровода мозга, а через 2 межжелудочковых отверстия, foramina interventriculares, - с боковыми желудочками полушарий большого мозга.
Подкорковые функции в механизмах формирования поведенческих реакций человека и животных функции подкорковых образований проявляются всегда в тесном взаимодействии с корой больших полушарий. К подкорковым образованиям относят структуры, лежащие между корой и продолговатым мозгом: таламус (см. Головной мозг), гипоталамус (см.), базальные узлы (см.), комплекс образований, объединяемых в лимбическую систему мозга, а также (см.) ствола мозга и таламуса. Последней принадлежит ведущая роль в формировании восходящих активирующих потоков возбуждения, генерализованно охватывающих кору больших полушарий. Любое афферентное возбуждение, возникшее при раздражении на периферии, на уровне ствола мозга трансформируется в два потока возбуждений. Один поток по специфическим путям достигает специфической для данного раздражения проекционной области коры; другой - от специфического пути по коллатералям попадает в ретикулярную формацию и от нее в виде мощного восходящего возбуждения направляется к коре больших полушарий, активируя ее (рис.). Лишенная связей с ретикулярной формацией кора головного мозга приходит в недеятельное состояние, характерное для состояния сна.
Схема восходящего активирующего влияния ретикулярной формации (по Мегуну): 1 и 2 - специфический (лемнисковый) проводящий путь; 3 - коллатерали, отходящие от специфического пути к ретикулярной формации ствола мозга; 4 - восходящая активирующая система ретикулярной формации; 5 - генерализованное влияние ретикулярной формации на кору больших полушарий.
Ретикулярная формация имеет тесные функциональные и анатомические связи с гипоталамусом, таламусом, продолговатым мозгом, лимбической системой, поэтому все наиболее общие функции организма (регуляция постоянства внутренней среды, дыхание, пищевая и болевая реакции) находятся в ее ведении. Ретикулярная формация является областью широкого взаимодействия потоков возбуждений различной природы, так как к ее нейронам конвергируют как афферентные возбуждения от периферических рецепторов (звуковых, световых, тактильных, температурных и др.), так и возбуждения, приходящие от других отделов головного мозга.
Афферентные потоки возбуждений от периферических рецепторов на пути к коре больших полушарий имеют многочисленные синаптические переключения в таламусе. От латеральной группы ядер таламуса (специфические ядра) возбуждения направляются по двум путям: к подкорковым ганглиям и к специфическим проекционным зонам коры мозга. Медиальная группа ядер таламуса (неспецифические ядра) служит местом переключения восходящих активирующих влияний, которые направляются от стволовой ретикулярной формации в кору мозга. Тесные функциональные взаимосвязи между специфическими и неспецифическими ядрами таламуса обеспечивают первичный анализ и синтез всех афферентных возбуждений, поступающих в головной мозг. У животных, находящихся на низких ступенях филогенетического развития, таламус и лимбические образования играют роль высшего центра интеграции поведения, обеспечивая все необходимые рефлекторные акты животного, направленные на сохранение его жизни. У высших животных и человека высшим центром интеграции является кора больших полушарий.
С функциональной точки зрения к подкорковым образованиям относят комплекс структур головного мозга, который играет ведущую роль в формировании основных врожденных рефлексов человека и животных: пищевых, половых и оборонительных. Этот комплекс получил название лимбической системы и включает в себя поясную извилину, гиппокамп, грушевидную извилину, обонятельный бугорок, миндалевидный комплекс и область перегородки. Центральное место среди образований лимбической системы отводится гиппокампу. Анатомически установлен гиппокампальный круг (гиппокамп → свод → мамиллярные тела → передние ядра таламуса → поясная извилина → cingulum → гиппокамп), который вместе с гипоталамусом играет ведущую роль в формировании . Регуляторные влияния лимбической системы широко распространяются на вегетативные функции (поддержание постоянства внутренней среды организма, регуляция кровяного давления, дыхания, сосудов, моторики желудочно-кишечного тракта, половых функций).
Кора больших полушарий оказывает постоянные нисходящие (тормозные и облегчающие) влияния на подкорковые структуры. Существуют различные формы циклического взаимодействия между корой и подкоркой, выражающиеся в циркуляции возбуждений между ними. Наиболее выраженная замкнутая циклическая связь существует между таламусом и соматосенсорной областью коры мозга, составляющими в функциональном отношении единое целое. Корково-подкорковая циркуляция возбуждений определяется не только таламокортикальными связями, но и более обширной системой подкорковых образований. На этом базируется вся условно-рефлекторная деятельность организма. Специфика циклических взаимодействий коры и подкорковых образований в процессе формирования поведенческой реакции организма определяется его биологическими состояниями (голод, боль, страх, ориентировочно - исследовательская реакция).
Подкорковые функции . Кора головного мозга является местом высшего анализа и синтеза всех афферентных возбуждений, областью формирования всех сложных приспособительных актов живого организма. Однако полноценная аналитико-синтетическая деятельность коры больших полушарий возможна лишь при условии прихода к ней от подкорковых структур мощных генерализованных потоков возбуждений, богатых энергией и способных обеспечить системный характер корковых очагов возбуждений. С этой точки зрения и следует рассматривать функции подкорковых образований, являющихся, по выражению , «источником энергии для коры».
В анатомическом плане к подкорковым образованиям относят нейрональные структуры, расположенные между корой головного мозга (см.) и продолговатым мозгом (см.), а с функциональной точки зрения - подкорковые структуры, которые в тесном взаимодействии с корой больших полушарий формируют целостные реакции организма. Таковы таламус (см.), гипоталамус (см.), базальные узлы (см.), так называемая лимбическая система мозга. С функциональной точки зрения к подкорковым образованиям относят и ретикулярную формацию (см.) ствола мозга и таламуса, которой принадлежит ведущая роль в формировании восходящих активирующих потоков к коре больших полушарий. Восходящие активирующие влияния ретикулярной формации открыли Моруцци и Мегун (G. Moruzzi, Н. W. Magoun). Раздражая электрическим током ретикулярную формацию, эти авторы наблюдали переход медленной электрической активности коры головного мозга в высокочастотную, низкоамплитудную. Такие же изменения электрической активности коры мозга («реакция пробуждения», «реакция десинхронизации») наблюдались при переходе от сонного состояния животного к бодрствующему. На основании этого возникло предположение о пробуждающем влиянии ретикулярной формации (рис. 1).

Рис. 1. «Реакция десинхронизации» корковой биоэлектрической активности при раздражении у кошки седалищного нерва (отмечено стрелками): СМ - сенсомоторная область коры мозга; ТЗ - теменно-затылочная область коры мозга (л - левая, п - правая).
В настоящее время известно, что реакция десинхронизации корковой электрической активности (активация коры мозга) может возникать при любом афферентном воздействии. Это связано с тем, что на уровне ствола мозга афферентное возбуждение, возникающее при раздражении любых рецепторов, трансформируется в два потока возбуждения. Один поток направляется по классическому лемнисковому пути и достигает специфической для данного раздражения корковой проекционной области; другой - попадает от лемнисковой системы по коллатералям в ретикулярную формацию и от нее в виде мощных восходящих потоков направляется к коре мозга, генерализованно активируя ее (рис. 2).

Рис. 2. Схема восходящего активирующего влияния ретикулярной формации (по Мегуну): 1-3 - специфический (лемнисковый) проводящий путь; 4 - коллатерали, отходящие от специфического пути к ретикулярной формации ствола мозга; 5 - восходящая активирующая система ретикулярной формации; в - генерализованное влияние ретикулярной формации на кору больших полушарий.
Это генерализованное восходящее активирующее влияние ретикулярной формации - непременное условие поддержания бодрствующего состояния мозга. Лишенная источника возбуждения, которым служит ретикулярная формация, кора головного мозга приходит в недеятельное состояние, сопровождаемое медленной высокоамплитудной электрической активностью, характерной для состояния сна. Такую картину можно наблюдать при децеребрации, т. е. у животного с перерезанным стволом мозга (см. ниже). В этих условиях ни какие-либо афферентные раздражения, ни прямое раздражение ретикулярной формации не вызывает диффузной, генерализованной реакции десинхронизации. Таким образом, доказано наличие в головном мозге по крайней мере двух основных каналов поступления афферентных воздействий на кору больших полушарий: по классическому лемнисковому пути и по коллатералям через ретикулярную формацию ствола мозга.
Так как при любом афферентном раздражении генерализованная активация коры мозга, оцениваемая по электроэнцефалографическому показателю (см. Электроэнцефалография), всегда сопровождается реакцией десинхронизации, многие исследователи пришли к выводу, что любые восходящие активирующие влияния ретикулярной формации на кору головного мозга являются неспецифическими. Главными аргументами в пользу такого вывода явились следующие: а) отсутствие сенсорной модальности, т. е. однотипность изменений биоэлектрической активности при воздействии различных сенсорных раздражителей; б) постоянный характер активации и генерализованное распространение возбуждения по коре, оцениваемое опять-таки по электроэнцефалографическому показателю (реакция десинхронизации). На этом основании все виды генерализованной десинхронизации корковой электрической активности признавались также едиными, не различающимися по каким-либо физиологическим качествам. Однако при формировании целостных приспособительных реакций организма восходящие активирующие влияния ретикулярной формации на кору головного мозга носят специфический характер, соответствующий данной биологической деятельности животного - пищевой, половой, оборонительной (П. К. Анохин). Это означает, что в формировании различных биологической реакций организма участвуют различные области ретикулярной формации, осуществляющие активацию коры больших полушарий (А. И. Шумилина, В. Г. Агафонов, В. Гавличек).
Наряду с восходящими влияниями на кору больших полушарий ретикулярная формация может оказывать и нисходящие влияния на рефлекторную деятельность спинного мозга (см.). В ретикулярной формации различают области, которые оказывают тормозящие и облегчающие влияния на моторную активность спинного мозга. По своему характеру эти влияния диффузны и оказывают воздействие на все группы мышц. Они передаются по нисходящим спинальным путям, которые различны для тормозящих и облегчающих влияний. О механизме ретикулоспинальных влияний существует две точки зрения: 1) ретикулярная формация оказывает тормозящие и облегчающие влияния непосредственно на мотоневроны спинного мозга; 2) эти влияния на мотоневроны передаются через клетки Реншо. Особенно четко нисходящие влияния ретикулярной формации выражены у децеребрированного животного. Децеребрация осуществляется путем перерезки головного мозга по передней границе четверохолмия. При этом развивается так называемая децеребрационная ригидность с резким увеличением тонуса всех мышц-разгибателей. Считают, что этот феномен развивается в результате перерыва путей, идущих от вышележащих образований мозга к тормозящему отделу ретикулярной формации, что обусловливает уменьшение тонуса этого отдела. В результате облегчающие влияния ретикулярной формации начинают преобладать, что и приводит к увеличению тонуса мышц.
Важной особенностью ретикулярной формации является ее высокая чувствительность к различным химическими веществам, циркулирующим в крови (СO 2 , адреналин и др.). Это обеспечивает включение ретикулярной формации в регулирование некоторых вегетативных функций. Ретикулярная формация является также местом избирательного действия многих фармакологических и медикаментозных препаратов, что используют при лечении некоторых заболеваний ЦНС. Высокая чувствительность ретикулярной формации к барбитуратам и ряду нейроплегических средств позволила по-новому представить механизм наркотического сна. Действуя тормозящим образом на нейроны ретикулярной формации, наркотик тем самым лишает кору головного мозга источника активирующих влияний и обусловливает развитие состояния сна. Гипотермическое действие аминазина и подобных ему препаратов объясняют влиянием этих веществ на ретикулярную формацию.
Ретикулярная формация имеет тесные функциональные и анатомические связи с гипоталамусом, таламусом, продолговатым мозгом и другими отделами головного мозга, поэтому все наиболее общие функции организма (терморегуляция, пищевая и болевая реакции, регуляция постоянства внутренней среды организма) находятся в той или иной функциональной зависимости от нее. Ряд исследований, сопровождавшийся регистрацией при помощи микроэлектродной техники электрической активности отдельных нейронов ретикулярной формации, показал, что эта область является местом взаимодействия афферентных потоков различной природы. К одному и тому же нейрону ретикулярной формации могут конвергировать возбуждения, возникающие не только при раздражении различных периферических рецепторов (звуковых, световых, тактильных, температурных и др.), но и идущие от коры больших полушарий, мозжечка и других подкорковых структур. На основе этого механизма конвергенции в ретикулярной формации происходит перераспределение афферентных возбуждений, после чего они в виде восходящих активирующих потоков направляются к нейронам коры головного мозга.
Прежде чем достигнуть коры, эти потоки возбуждения имеют многочисленные синаптические переключения в таламусе, который служит как бы промежуточным, связующим звеном между низшими образованиями ствола мозга и корой больших полушарий. Импульсы от периферических концов всех внешних и внутренних анализаторов (см.) переключаются в латеральной группе ядер таламуса (специфические ядра) и отсюда направляются по двум путям: к подкорковым ганглиям и к специфическим проекционным зонам коры мозга. Медиальная группа ядер таламуса (неспецифические ядра) служит местом переключения восходящих активирующих влияний, которые направляются от стволовой ретикулярной формации в кору мозга.
Специфические и неспецифические ядра таламуса находятся в тесной функциональной взаимосвязи, что обеспечивает первичный анализ и синтез всех афферентных возбуждений, поступающих в головной мозг. В таламусе имеется четкая локализация представительства различных афферентных нервов, идущих от различных рецепторов. Эти афферентные нервы заканчиваются в определенных специфических ядрах таламуса, а от каждого ядра волокна направляются в кору головного мозга к специфическим проекционным зонам представительства той или иной афферентной функции (зрительной, слуховой, тактильной и т. д.). Особенно тесно таламус связан с соматосенсорной областью коры больших полушарий. Эта взаимосвязь осуществляется благодаря наличию замкнутых циклических связей, направленных как от коры к таламусу, так и от таламуса к коре. Поэтому соматосенсорную область коры и таламус в функциональном отношении можно рассматривать как единое целое.
У животных, находящихся на более низких ступенях филогенетического развития, таламус играет роль высшего центра интеграции поведения, обеспечивая все необходимые рефлекторные акты животного, направленные на сохранение его жизни. У животных, стоящих на высших ступенях филогенетической лестницы, и у человека высшим центром интеграции становится кора больших полушарий. Функции же таламуса заключаются в регуляции и осуществлении ряда сложных рефлекторных актов, являющихся как бы базой, на основе которой создается адекватное целенаправленное поведение животного и человека. Эти ограниченные функции таламуса четко проявляются у так называемого таламического животного, т. е. у животного с удаленными корой больших полушарий и подкорковыми узлами. Такое животное может самостоятельно передвигаться, сохраняет основные позно-тонические рефлексы, обеспечивающие нормальное положение тела и головы в пространстве, сохраняет регуляцию температуры тела и всех вегетативных функций. Но оно не может адекватно реагировать на различные раздражители внешней среды вследствие резкого нарушения условно-рефлекторной деятельности. Таким образом, таламус в функциональной взаимосвязи с ретикулярной формацией, оказывая локальные и генерализованное воздействия на кору больших полушарий, организует и регулирует соматическую функцию головного мозга как целого.
Среди структур головного мозга, относящихся к подкорковым с функциональной точки зрения, выделяют комплекс образований, который играет ведущую роль в формировании основных врожденных активностей животного: пищевой, половой и оборонительный. Этот комплекс получил название лимбической системы мозга и включает в себя гиппокамп, грушевидную извилину, обонятельный бугорок, миндалевидный комплекс и область перегородки (рис. 3). Все эти образования объединяются на функциональной основе, так как они принимают участие в обеспечении поддержания постоянства внутренней среды, регуляции вегетативных функций, в формировании эмоций (см.) и мотиваций (см.). Многие исследователи относят к лимбической системе и гипоталамус. Лимбическая система принимает непосредственное участие в формировании эмоционально окрашенных, примитивных врожденных форм поведения. Особенно это относится к формированию половой функции. При поражении (опухоль, травма и др.) некоторых структур лимбической системы (височная область, поясная извилина) у человека нередко наблюдаются сексуальные расстройства.

Рис. 3. Схематическое изображение основных связей лимбической системы (по Мак-Лейну): N - nucleus interpeduncularis; MS и LS - медиальная и латеральная обонятельные полоски; S - перегородка; MF - медиальный пучок переднего мозга; Т - обонятельный бугорок; AT - переднее ядро таламуса; М - мамиллярное тело; SM - stria medialis (стрелками обозначено распространение возбуждения по лимбической системе).
Центральное место среди образований лимбической системы отводится гиппокампу. Анатомически установлен гиппокампальный круг (гиппокамп → свод → мамиллярные тела → передние ядра таламуса → поясная извилина → cingulum → гиппокамп), который вместе с, гипоталамусом (си.) играет ведущую роль в формировании эмоций. Непрерывная циркуляция возбуждения по гиппокампальному кругу определяет главным образом тоническую активацию коры головного мозга, а также интенсивность эмоций.
Часто у больных с тяжелыми формами психоза и другими психическими заболеваниями после смерти находили патологические изменения в структурах гиппокампа. Предполагают, что циркуляция возбуждения по гиппокампальному кольцу служит одним из механизмов памяти. Отличительная особенность лимбической системы - тесная функциональная взаимосвязь между ее структурами. Благодаря этому возбуждение, возникшее в какой-либо структуре лимбической системы, тут же охватывает остальные образования и долгое время не выходит за пределы всей системы. Подобное длительное, «застойное» возбуждение лимбических структур, вероятно, также лежит в основе формирования эмоциональных и мотивационных состояний организма. Некоторые образования лимбической системы (миндалевидный комплекс) оказывают генерализованное восходящее активирующее влияние на кору головного мозга.
Учитывая регуляторные влияния лимбической системы на вегетативные функции (кровяное давление, дыхание, тонус сосудов, моторику желудочно-кишечного тракта), можно понять те вегетативные реакции, которые сопровождают любой условнорефлекторный акт организма. Этот акт как целостная реакция осуществляется всегда при непосредственном участии коры больших полушарий, которая является высшей инстанцией анализа и синтеза афферентных возбуждений. У животных после удаления коры головного мозга (декортицированных) резко нарушается условно-рефлекторная деятельность, причем, чем выше стоит животное в эволюционном отношении, тем ярче выражены эти нарушения. Поведенческие реакции животного, подвергшегося декортикации, сильно расстраиваются; большую часть времени такие животные спят, просыпаясь только при сильных раздражениях и для совершения простых рефлекторных актов (мочеиспускание, дефекация). У таких животных можно выработать условнорефлекторные реакции, однако слишком примитивные и недостаточные для осуществления адекватной приспособительной деятельности организма.
Вопрос о том, на каком уровне головного мозга (в коре или подкорке) происходит замыкание условного рефлекса, в настоящее время не рассматривается как принципиальный. Мозг участвует в формировании приспособительного поведения животного, в основе которого лежит принцип условного рефлекса, как единая целостная система. Любые раздражители - как условные, так и безусловные - конвергируют к одному и тому же нейрону различных подкорковых образований, а также к одному нейрону различных областей коры больших полушарий. Изучение механизмов взаимодействия коры и подкорковых образований в процессе формирования поведенческой реакции организма - одна из основных задач современной физиологии головного мозга. Кора больших полушарий, являясь высшей инстанцией синтеза афферентных возбуждений, организует внутренние нервные связи для совершения ответного рефлекторного акта. Ретикулярная формация и другие подкорковые структуры, оказывая множественные восходящие влияния на кору головного мозга, создают лишь необходимые условия для организации более совершенных корковых временных связей, а в результате этого - и для формирования адекватной поведенческой реакции организма. Кора больших полушарий в свою очередь оказывает постоянные нисходящие (тормозные и облегчающие) влияния на подкорковые структуры. В этом тесном функциональном взаимодействии между корой и нижележащими образованиями головного мозга заключена основа интегративной деятельности мозга как единого целого. С этой точки зрения, разделение функций мозга на чисто корковые и чисто подкорковые в какой-то степени искусственно и необходимо лишь для понимания роли различных образований мозга в формировании целостной приспособительной реакции организма.
Регуляция вегетативных функций может быть связана с их гормональной регуляцией
H а п p и м е p в о з б у ж д е н и е СИМПАТИЧЕСКОГО отдела увеличивает секрецию адреналина и норадреналина в мозговом слое надпочечников.Следствием является повышение сахара в крови ПАРАСИМПАТИЧЕСКОГО - стимулирует выход инсулина в островках Лангерганса поджелудочной железы, следствием чего является отложение сахара в печени в форме гликогена
Таким образом, центральная регуляция вегетативных функций о с у щ е с т в л я е т с я сложным многоэтажным комплексом центров и многокомпонентными нервными образованиями с участием гормональных механизмов.
Высшими центрами вегетативной регуляции являются
ПЕРЕДНИЙ ГИПОТАЛАМУС трофотропные центры (парасимпатические)
ЗАДНИЙ ГИПОТАЛАМУС эрготрофные центры (симпатические)
СРЕДНИЙ ГИПОТАЛАМУС центры обмена веществ и терморегуляции
ЛИМБИЧЕСКАЯ СИСТЕМА центры интеграции вегетативных и соматических функций, эмоциональных реакций
КОРА ГОЛОВНОГО МОЗГА центры условных рефлексов, организации поведения
Гипоталамус находится на дне и по бокам III желудочка мозга. Структуры: серый бугор, воронка, сосцевидные тела. Зоны: гипофизотропная (преоптические и передние ядра), медиальная (средние ядра), латеральная (наружные, задние ядра).Физиологическая роль – высший подкорковый интегративный центр вегетативной нервной системы, который оказывает действие на:
1) терморегуляцию. Передние ядра – это центр теплоотдачи, где происходит регуляция процесса потоотделения, частоты дыхания и тонуса сосудов в ответ на повышение температуры окружающей среды. Задние ядра – центр теплопродукции и обеспечения сохранности тепла при понижении температуры;
2) гипофиз. Либерины способствуют секреции гормонов передней доли гипофиза, статины тормозят ее;
3) жировой обмен. Раздражение латеральных (центра питания) ядер и вентромедиальных (центра насыщения) ядер ведет к ожирению, торможение – к кахексии;
4) углеводный обмен. Раздражение передних ядер ведет к гипогликемии, задних – к гипергликемии;
5) сердечно-сосудистую систему. Раздражение передних ядер оказывает тормозное влияние, задних – активирующее;
6) моторную и секреторную функции ЖКТ. Раздражение передних ядер повышает моторику и секреторную функцию ЖКТ, задних – тормозит половую функцию. Разрушение ядер ведет к нарушению овуляции, сперматогенеза, снижению половой функции;
7) поведенческие реакции. Раздражение стартовой эмоциональной зоны (передних ядер) вызывает чувство радости, удовлетворения, эротические чувства, стопорной зоны (задних ядер) вызывает страх, чувство гнева, ярости.
37. Условный рефлекс как форма приспособления человека к изменяющимся условиям существования. Отличия условных и безусловных рефлексов. Закономерности образования и проявления условных рефлексов.
Условные рефлексы вырабатываются в течение жизни, так как не имеют готовых рефлекторных дуг. Они носят индивидуальный характер и в зависимости от условий существования могут постоянно меняться. Их особенности:
морфологическим субстратом является кора больших полушарий, при ее удалении старые рефлексы исчезают, а новые не вырабатываются;
на их базе формируется взаимодействие организма с внешней средой, т. е. они уточняют, усложняют и делают тонкими данные отношения.
Итак, условные рефлексы – это приобретенный в течение жизни набор поведенческих реакций. Их классификация:
по природе условного раздражителя выделяют натуральные и искусственные рефлексы. Натуральные рефлексы вырабатываются на естественные качества раздражителя (например, вид пищи), а искусственные – на любые;
по рецепторному признаку – экстероцептивные, интероцептивные и проприоцептивные;
в зависимости от структуры условного раздражителя – простые и сложные;
по эфферентному пути – соматические (двигательные) и вегетативные (симпатические и парасимпатические);
по биологическому значению – витальные (пищевые, оборонительные, локомоторные), зоосоциальные, ориентировочные;
по характеру подкрепления – низшего и высшего порядка;
в зависимости от сочетания условного и безусловного раздражителя – наличные и следовые.
Таким образом, условные рефлексы вырабатываются в течение жизни и имеют большое значение для человека.
Для образования условных рефлексов необходимы определенные условия.
1. Наличие двух раздражителей – индифферентного и безусловного. Это связано с тем, что адекватный раздражитель вызовет безусловный рефлекс, а уже на его базе будет вырабатываться условный. Индифферентный раздражитель гасит ориентировочный рефлекс.
2. Определенное сочетание во времени двух раздражителей. Сначала должен включиться индифферентный, а затем безусловный, причем промежуточное время должно быть постоянным.
3. Определенное сочетание по силе двух раздражителей. Индифферентный – пороговой, а безусловный – сверхпороговой.
4. Полноценность ЦНС.
5. Отсутствие посторонних раздражителей.
6. Многократное повторение действия раздражителей для возникновения доминантного очага возбуждения.
В экспериментальных условиях доказано, что образование условного рефлекса происходит в три этапа:
1) знакомство;
2) выработка условного рефлекса, после погашения действия ориентировочного;
3) закрепление выработанного условного рефлекса.
Закрепление происходит в две стадии. Вначале условный рефлекс возникает и на действие похожих раздражителей из-за иррадиации возбуждения. Через небольшой промежуток времени уже только на условный сигнал, так как происходит концентрация процессов возбуждения в области проекции в коре больших полушарий.
Центры вегетативной нервной системы разделяют на сегментарные и надсегментарные (высшие вегетативные центры).
Сегментарные центры располагаются в нескольких отделах центральной нервной системы, где выделяют 4 очага:
1. Мезенцефалический отдел в среднем мозге - добавочное ядро (Якубовича), nucleus accessorius, и непарное срединное ядро глазодвигательного нерва (III пара).
2. Бульбарный отдел в продолговатом мозге и мосту - верхнее слюноотделительное ядро, nucleus salivatorius superior, промежуточно-лицевого нерва (VII пара), нижнее слюноотделительное ядро, nucleus salivatorius inferior, языкоглоточного нерва (IX пара) и дорсальное ядроблуждающего нерва (X пара), nucleus dorsalis n. vagi.
Оба этих отдела объединяются под названием краниального и относятся к парасимпатическим центрам.
3. Тораколюмбальный отдел - промежуточно-боковые ядра, nuclei intermediolaterales, 16-ти сегментов спинного мозга (С8, Th1-12, L1-3). Они являются симпатическими центрами.
4. Сакральный отдел - промежуточно-боковые ядра, nuclei intermediolaterales, 3-х крестцовых сегментов спинного мозга (S2-4) и относятся к парасимпатическим центрам.
Высшие вегетативные центры (надсегментарные) объединяют и регулируют деятельность симпатического и парасимпатического отделов, к ним относятся:
1. Ретикулярная формация, ядра которой формируют центры жизненно-важных функций (дыхательный и сосудодвигательный центры, центры сердечной деятельности, регуляции обмена веществ и т.д.). Проекция дыхательного центра соответствует средней трети продолговатого мозга, сосудодвигательного центра - нижней части ромбовидной ямки. Нарушение функции ретикулярной формации проявляется вегетативно-сосудистыми расстройствами (кардио-васкулярные, вазомоторные). Кроме того страдают интегративные функции, которые необходимы для формирования целесообразного адаптивного поведения.
2. Мозжечок, принимая участие в регуляции двигательных актов, одновременно обеспечивает эти анимальные функции адаптационно-трофическими влияниями, которые через соответствующие центры приводят к расширению сосудов интенсивно работающих мышц, повышению уровня трофических процессов в последних. Установлено участие мозжечка в регуляции таких вегетативных функций, как зрачковый рефлекс, трофика кожи (скорость заживления ран), сокращение мышц, поднимающих волосы.
3. Гипоталамус - главный подкорковый центр интеграции вегетативных функций, имеет существенное значение в поддержании оптимального уровня обмена веществ (белкового, углеводного, жирового, минерального, водного) и терморегуляции. За счет связей с таламусом он получает разностороннюю информацию о состоянии органов и систем организма, а вместе с гипофизом образует функциональный комплекс - гипоталамо-гипофизарную систему. Гипоталамус в ней выполняет роль своеобразного реле, включающего гипофизарную гормональную цепь в регуляцию различных висцеральных и соматических функций.
4. Особое место занимает лимбическая система обеспечивающая интеграцию вегетативных, соматических и эмоциональных реакций.
5. Полосатое тело имеет ближайшее отношение к безусловнорефлекторной регуляции вегетативных функций. Повреждение или раздражение ядер полосатого тела вызывает изменение кровяного давления, усиление слюно- и слезоотделения, усиление потоотделения.
Высшим центром регуляции вегетативных и соматических функций, а также их координации является кора полушарий большого мозга. Непрерывный поток импульсов от органов чувств, сомы и внутренних органов по афферентным путям поступает в кору головного мозга и через эфферентную часть вегетативной нервной системы, главным образом через гипоталамус, кора оказывает соответствующее влияние на функцию внутренних органов, обеспечивая адаптацию организма к меняющимся условиям окружающей и внутренней среды. Примером кортиковисцеральной связи может служить изменение вегетативных реакций под влиянием словесных сигналов (через вторую сигнальную систему).
Таким образом, вегетативная нервная система, так же как и вся нервная система, построена по принципу иерархии, подчиненности. Схему организации вегетативной иннервации иллюстрирует рис.1.
Рис. 1 Принцип организации вегетативной нервной системы.
Мозжечок координирует сложные двигательные акты и произвольные движения. Эфферентные влияния мозжечка через верхние ножки направляются к красному ядру среднего мозга, к ядрам таламуса и гипоталамуса, к подкорковым узлам и к двигательной зоне коры больших полушарий. Через красноядерно-спинальный путь мозжечок регулирует деятельность мотонейронов спинного мозга.
Афферентные импульсы поступают в мозжечок через нижние и средние ножки. По спинно-мозжечковым путям Флексига и Говерса в мозжечок поступают импульсы от проприоцепторов двигательного аппарата. Из ядер Голя и Бурдаха продолговатого мозга к мозжечку поступают проприоцептивные сигналы, направляющиеся к корковым проекциям. Через средние ножки мозжечок связан с вестибулярными ядрами и двигательными зонами коры больших полушарий.
Многочисленные связи мозжечка со всеми отделами центральной нервной системы способствуют выполнению им сложной интегративной функции по координации движений. Нарушение целостности мозжечка приводит к расстройствам движений, нарушению пространственной ориентировки, потере тонуса. Мышцы теряют способность к слитным, целостным движениям. Голова и туловище производят качательные и дрожательные движения (астазия). Точность и пространственная упорядоченность движений нарушаются (дисметрия), координация при выполнении сложных движений утрачивается (атаксия). Теряется способность сохранять равновесие (дезэквилибрия).
Наряду с двигательными расстройствами отмечается повышенная утомляемость (астения) и падение мышечного тонуса (атония). Произвольная двигательная деятельность регулируется мозжечком через двусторонние связи с корковыми отделами двигательного анализатора. Через ретикулярную формацию и γ-мотонейроны спинного мозга мозжечок контролирует мышечный тонус. Явления астазии объясняются снятием тормозных влияний мозжечка на проприоцептивные рефлексы.
Роль мозжечка в координации движений проявляется в согласовании быстрых и медленных компонентов двигательных актов. Постоянная текущая коррекция движения делает его слитным, строго приуроченным к определенным условиям.
«Физиология человека», Н.А. Фомин
Кортикоретикулярные связи: А — схема путей восходящих активирующих влияний; Б — схема нисходящих влияний коры; Сn — специфические афферентные пути к коре с коллатералями к ретикулярной формации (по Мэгуну). Головной и спинной мозг осуществляют две формы регуляторных влияний: специфическую и неспецифическую. К специфической системе регуляции относятся нервные пути, проводящие эфферентные импульсы от всех рецепторов, центры…
Ретикулярная формация повышает возбудимость у мотонейронов спинного мозга, регулирующих активность мышечных веретен. Вследствие этого мышечные веретена посылают постоянный поток импульсов к спинному мозгу и возбуждают α-мотонейроны. В свою очередь, поток импульсов с α-мотонейронов поддерживает постоянный тонус скелетных мышц. Регуляторные тонические влияния поступают от покрышки мозга по двум путям ретикулоспинального тракта, проводящим нервные сигналы с разной…
К подкорковым ядрам относятся хвостатое ядро, бледный шар и скорлупа. Они находятся в толще больших полушарий головного мозга, между лобными долями и промежуточным мозгом. Эмбриональное происхождение хвостатого ядра и скорлупы едино, поэтому о них говорят иногда как о едином — полосатом теле (стриатум). Бледный шар, филогенетически наиболее древнее образование, обособлен от полосатого тела и морфологически,…
Раздражение бледного шара вызывает медленные тонические сокращения скелетных мышц. Бледный шар выполняет роль коллектора, связывающего полосатое тело с ядрами гипоталамуса, ствола мозга и таламуса. Важная роль принадлежит бледному шару и в регуляции гемодинамики. Разрушение полосатого тела вызывает у животных снижение чувствительности к тактильным и болевым раздражителям. Теряются ориентировочные рефлексы, появляется «эмоциональная тупость». Нарушаются процессы запоминания:…
Передние отделы новой коры ориентируют поведение на сигналы, обладающие высокой вероятностью подкрепления, иначе говоря, удовлетворения потребностей. На сигналы с малой вероятностью подкрепления отвечает гиппокамп. В случае ослабления коркового контроля возможна субъективная оценка маловероятных событий, как событий с большой вероятностью подкрепления. События с принципиально различной вероятностью подкрепления представляются равновероятными. Разгоряченному мозгу гоголевского Хлестакова тридцать, пять тысяч…
Рекомендуем также
 Как приготовить джем из ирги: рецепты вкусного ягодного джема
Как приготовить джем из ирги: рецепты вкусного ягодного джема
 Употребление времен в английском языке: логика использования В настоящее время он наиболее
Употребление времен в английском языке: логика использования В настоящее время он наиболее
 Самая близкая к земле планета Две самые близкие планеты к земле
Самая близкая к земле планета Две самые близкие планеты к земле
 Вестник богов в греческой мифологии
Вестник богов в греческой мифологии
 Хлор очень сильный окислитель
Хлор очень сильный окислитель
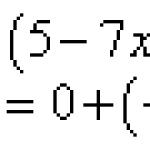 Производная произведения двух функций
Производная произведения двух функций